 2022, Vol. 34
2022, Vol. 34 


(2: 南京林业大学生物与环境学院, 南京 210037)
(2: College of Biology and the Environment, Nanjing Forestry University, Nanjing 210037, P. R. China)
湿地生态系统发挥着不可替代的生态系统服务功能[1-2]. 人类利用湿地的历史由来已久,其丰富的自然资源可为人类社会的发展提供必要的生产、生活资料. 同时,湿地也是众多生物的栖息、繁殖地,对维持全球生物多样性具有重要意义[3]. 然而,随着人类活动的加剧,全球湿地资源丧失严重,破碎化程度日益升高,产生了一系列的生态后果,最终可能导致生态系统服务功能的降低乃至丧失[4-5]. 因此,保护湿地资源、维持湿地生态系统健康是人类社会面临的重大环境问题之一,受到了全世界的广泛关注.
水鸟是湿地生态系统中主要生物类群之一,因其具有种类丰富、便于观察等特点,常被作为湿地健康状况的重要指示物种[6]. 水鸟的丰富度和多样性受众多因素制约,根据岛屿生物地理学理论,较大的湿地可能有更高的生物多样性[7]. 湿地内部的栖息地状况是决定越冬水鸟丰富度和多样性的重要因素. 然而,鸟类有极强的飞行能力,在湿地内部食物短缺时可以利用周围的适宜栖息地. 湿地周围的农田、植被等土地利用类型也可能影响越冬水鸟多样性[8],如部分水鸟可能利用湿地周边的农田或植被作为其觅食地或栖息地[9],因此,湖泊湿地周围栖息地状况也是影响其越冬水鸟丰富度和多样性的重要因素.
近年来,随着人类活动的日益加剧,人为因素成为了影响水鸟分布和多样性最主要的因素[10-11]. 人类活动可以直接影响鸟类的丰富度,如人类活动产生的“恐惧景观”可以降低鸟类对栖息地的利用率[12]. 不透水表面(如房屋、道路等)通常被作为衡量人为干扰的重要指标,有研究表明,栖息地距最近房屋的距离与水鸟丰富度和多样性呈正相关[10]. 道路可以提高湿地的可达性,导致人为干扰的增加,进而对水鸟多样性产生负面影响[13]. 人类对栖息地景观的改造极大地影响了栖息地的适合度,如导致栖息地丧失、破碎化等问题,进而影响越冬候鸟的丰富度和多样性[14-15]. 人均GDP也常被用作衡量人为干扰的重要指标,人均GDP较高的地区对栖息地的干扰程度也可能较大,进而导致鸟类多样性降低[16].
我国长江中下游地区气候温和、湖网密布,形成了世界独特的江-河复合型生态系统. 周期性的水位落差为越冬候鸟提供了良好的越冬场所,是东亚-澳大利西亚迁徙通道上最重要的水鸟越冬地和迁徙停歇地之一,每年有超过百万只水鸟在此越冬,对维持全球水鸟多样性有重要意义[17-18]. 然而,该地区也是我国经济最发达的区域之一,人类活动及其导致的栖息地变化已经对区域越冬水鸟多样性造成重要威胁. 研究表明,栖息地的丧失和退化是东亚-澳大利西亚迁徙鸟类所面临的主要威胁[19]. 在东洞庭湖地区的相关研究也表明,栖息地丧失和破碎化导致了该地区越冬鹤类种群数量的持续下降[20].
目前,对长江中下游地区越冬水鸟多样性的研究多集中在部分国家级保护区或国际重要湿地,如江西鄱阳湖、湖南洞庭湖和安徽升金湖等. 在这些湖泊湿地中,水文环境的周期性变化形成了多样化的湿地景观类型,如草洲、泥滩等,为多种鸟类提供了适宜的越冬场所[11]. 鉴于当前区域发展水平的差异和对未来发展的预期,分析较发达的江苏沿江地区湖泊湿地环境和人为因素对水鸟多样性的影响,进而制定针对性的保护策略,不仅可为该地区越冬水鸟及其栖息地保护提供科学依据,也为流域欠发达地区寻求“环境保护新道路”提供借鉴. 本研究选择10处江苏沿江代表性湖泊湿地,基于2018-2020年越冬水鸟调查数据,分析环境和人为因素对区域越冬候鸟多样性的影响,并提出相应保护策略,为区域生物多样性保护提供参考.
1 研究区域和方法 1.1 研究区域江苏沿江地区位于亚洲大陆东岸中纬度地带,属亚热带湿润季风气候,四季分明,雨量充沛,境内河川交错,水网密布,长江横穿东西,河湖湿地众多,是我国湖泊分布较多的地区之一,尤其以东部太湖平原的河网最为密集,包括太湖、阳澄湖、滆湖等大型湖泊. 本研究选择江苏沿江地区10处代表性湖泊湿地作为研究区域,开展越冬水鸟多样性调查. 这些湖泊均为自然湿地,是江苏沿江地区水鸟丰富度和多样性较高的区域,分布在江苏省沿江各行政区域,且经济发展具有一定的差异. 同时,所选湖泊也考虑了面积梯度,以便后续分析,相关区位等信息如表 1.
| 表 1 江苏沿江地区10处代表性湖泊湿地区位及水域面积 Tab. 1 Geographical locations and areas of the selected 10 typical coastal lake wetlands along the Yangtze River in Jiangsu Province |
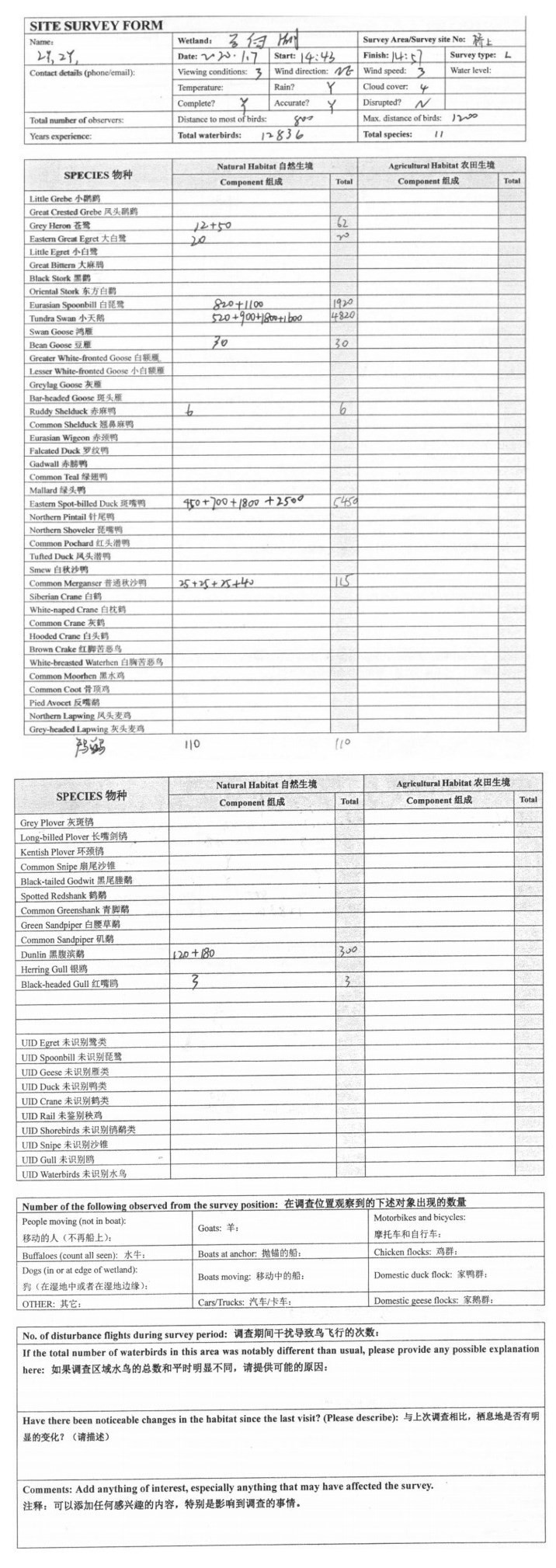
2018-2020年,在每年越冬水鸟群落最稳定的月份(1月),对所选10处代表性湖泊湿地开展1次越冬水鸟多样性调查. 调查采用样点法进行分区计数,根据湿地大小、地形、地貌等特征设置不同数量的调查位点,位点设置以尽量覆盖整个湿地范围为原则,调查位点分布如图 1,每个湿地调查位点设置详情见附图Ⅰ. 调查时,调查人员分为两组,借助20×60单双筒望远镜(Swarovski,奥地利)和10×42双筒望远镜(胜图,中国)对每个调查位点逐一开展调查工作,记录观察范围内所有水鸟的种类和数量. 水鸟数量较少时采用直接计数法,对于较大的水鸟集群则采用集团计数法. 鸟类分类系统依据《中国鸟类分类与分布名录》(第三版)[21]. 调查中同时记录调查人员信息、天气情况、能见度等信息(调查表格样表见附表Ⅰ),并对每个调查位点拍摄东、西、南、北4个方位的环境照片.

|
图 1 江苏沿江10处代表性湖泊湿地分布及调查位点设置 Fig.1 Distribution of the 10 typical lake wetlands of the Yangtze River coast in Jiangsu Province and the survey points design |
基于2018-2020年水鸟调查数据,将每处湖泊湿地历次调查中某一物种的最大数量作为该湿地中此物种的最终数量,按如下方法,分别计算每处湖泊湿地水鸟群落的α和β多样性指数.
1)α多样性指数选择香农-威纳(Shannon-Wiener,H′)和逆辛普森(Inverse-Simpson,D)多样性指数[22],计算公式如下:
| $ {H^\prime } = - \sum\limits_{i = 1}^s {{P_i}} \ln {P_i} $ | (1) |
| $ D = \frac{1}{{\sum\limits_{i = 1}^s {P_i^2} }} $ | (2) |
式中,S为每处湖泊湿地水鸟的物种数,Pi是第i种水鸟的数量占所有水鸟数量的比值.
2) β多样性分析不同群落间物种组成的差异,由物种周转或替换和嵌套两种过程决定. 物种周转或替换表示不同群落间物种替换,而嵌套的产生则是因为物种丧失而导致的物种数的变化,拥有较少物种的群落成为拥有较多物种的群落的子集. β多样性指数选取Sørenson(βsor)成对相异性指数,并将其分解为物种空间转化组分(βsim)和嵌套组分(βsne),其计算公式如下[23]:
| $ \beta_{\mathrm{sor}}=\frac{b+c}{2 a+b+c} $ | (3) |
| $ \beta_{\mathrm{sim}}=\frac{\min (b, c)}{a+\min (b, c)} $ | (4) |
| $ \beta_{\mathrm{sne}}=\frac{|b-c|}{2 a+b+c} \times \frac{a}{a+\min (b, c)} $ | (5) |
式中,a为2个湖泊湿地共有的物种数,b和c分别是2个湖泊湿地各自特有的物种数.
总体β多样性指数选用Sørensen多点相异性指数,并参考上述Sørensen成对相异性指数的方法将其分解为空间周转(βSIM)和嵌套组分(βSNE),以探讨这两种过程如何影响物种分布格局. 使用各个组分所占的比例表示对总体β多样性的贡献,即βratio =βSNE/βSOR. 当βratio < 0.5则表示β多样性主要由物种空间周转决定;反之,则表示β多样性主要由嵌套组分决定[23-24]. 计算公式如下:
| $ \beta_{\mathrm{SOR}}=\frac{\left[\sum_{i<j} \min \left(b_{i j}, b_{j i}\right)\right]+\left[\sum_{i <j} \max \left(b_{i j}, b_{j i}\right)\right]}{2\left[\sum_i S_i-S_{\mathrm{T}}\right]+\left[\sum_{i<j} \min \left(b_{i j}, b_{j i}\right)\right]+\left[\sum_{i<j} \max \left(b_{i j}, b_{j i}\right)\right]} $ | (6) |
| $ \beta_{\mathrm{SIM}}=\frac{\left[\sum_{i<j} \min \left(b_{i j}, b_{j i}\right)\right]}{\left[\sum_i S_i-S_{\mathrm{T}}\right]+\left[\sum_{i<j} \min \left(b_{i j}, b_{j i}\right)\right]} $ | (7) |
| $ \begin{gathered} \beta_{\mathrm{SNE}}=\frac{\left[\sum_{i<j} \max \left(b_{i j}, b_{j i}\right)\right]-\left[\sum_{i<j} \min \left(b_{i j}, b_{j i}\right)\right]}{2\left[\sum_i S_i-S_{\mathrm{T}}\right]+\left[\sum_{i<j} \min \left(b_{i j}, b_{j i}\right)\right]+\left[\sum_{i<j} \max \left(b_{i j}, b_{j i}\right)\right]} \times \\ \frac{\left[\sum_i S_i-S_{\mathrm{T}}\right]}{\left[\sum_i S_i-S_{\mathrm{T}}\right]+\left[\sum_{i<j} \min \left(b_{i j}, b_{j i}\right)\right]} \end{gathered} $ | (8) |
式中,bij为湖泊湿地i中的物种数,bji为湖泊湿地j中的物种数,Si为湖泊湿地i中的物种总数,ST为所有湖泊湿地的总物种数.
1.2.3 变量越冬水鸟的丰富度和多样性可能受湿地本身及周边土地利用情况的影响. 因此,基于Landsat 8 OLI_TIRS数字卫星影像数据(拍摄时间:2019年1月23日;分辨率:30 m×30 m),我们对每个湖泊湿地划定了2 km范围的缓冲区,利用改进的归一化水体指数(MWDMI)提取水面部分. 由于缓冲区农田和植被在夏季影像中光谱相似,因此使用冬季影像,结合Google Earth地形图对农田、植被和不透水表面进行分类. 由于缓冲区内水体面积较小,主要为鱼塘和河流,且冬季水生植被很少,基于遥感影像进行植被分类难度极大,故没有进行单独提取. 以上所有处理均基于ENVI 5.3软件. 最后利用ArcGIS数据分析工具中的缓冲区分析等分析模块,计算缓冲区范围内的农田、植被、水面和不透水表面等的面积.
因此,我们通过收集、查阅湿地所在县(区)2019年政府年度工作报告获得人均GDP数据,如湿地隶属于多个县(区)级行政区,则计算其平均值代替.
1.2.4 数据分析利用R语言中vegan程序包计算每个湖泊湿地的香农-威纳和逆辛普森多样性指数. 利用R语言adespatial程序包中beta.div.comp函数计算单个湖泊湿地对区域β多样性的相对贡献(local contributions to beta diversity,LCBD index),相对贡献较大的湖泊湿地表明其可能具有较高的保护价值[25].
对湖泊湿地总面积、缓冲区植被、农田、水面以及不透水表面面积进行对数转换,然后进行线性模型分析. 模型分析前,先采用皮尔逊(Pearson)相关性分析检验所有自变量间的两两相关性(数据符合正态分布),对于相关性较高的成对变量(|r|>0.6),则去除其中一个以避免后续线性模型分析中的多重共线性问题. 经过该分析后,因农田面积与缓冲区植被面积(rp=0.76, P=0.01)以及不透水表面面积与人均GDP(rp=0.68, P=0.03)显著相关,因我们更关注植被面积和人均GDP对水鸟多样性的影响,因此将农田面积和不透水表面面积移除.
计数型数据类型通常符合泊松分布,因此,我们利用基于泊松分布的广义线性模型分析环境因素对水鸟丰富度和物种数的影响. 因变量分别为水鸟数量和物种数,自变量为湖泊湿地总面积,缓冲区植被、农田面积和人均GDP;利用多重线性模型分析环境因素对香农-威纳和逆辛普森多样性的影响.
所有数据分析在RStudio中加载vegan、betapart、adespatial、lme4等程序包完成.
2 研究结果 2.1 水鸟组成2018-2020年调查中,江苏沿江10处代表性湖泊湿地共记录到越冬水鸟42种(详见附表Ⅱ),其中2018年记录到23种共20751只;2019年记录到27种共45093只;2020年记录到32种共44154只. 按地理区系划分,古北界16种,广布种15种,古北界-东洋界9种,东洋界2种;按生态类群划分,有游禽22种,涉禽20种. 42种水鸟隶属于6目9科. 鹤形目(Gruiformes)和目(Podicipediformes)均为1科2种,占科、种总数的比例分别为11.1 % 和4.8 %;鹈形目(Pelecaniformes)为1科1种,占科、种总数的比例分别为11.1 % 和2.4 %;鹳形目(Ciconiiformes)为2科7种,占科、种总数的比例分别为22.2 % 和16.7 %;鸻形目(Charadriiformes)3科11种,其科、种数分别占总数的33.3 % 和26.2 %;雁形目(Anseriformes)水鸟种类最多,共计1科19种,占科、种总数的比例分别为11.1 % 和45.2 %.
记录到的水鸟中有国家Ⅰ级重点保护鸟类1种,为东方白鹳(Ciconia boyciana);国家Ⅱ级重点保护鸟类4种,分别为白琵鹭(Platalea leucorodia)、小天鹅(Cygnus columbianus)、白额雁(Anser albifrons)和斑头秋沙鸭(Mergellus albellus). 全球受胁物种有东方白鹳(濒危,EN)和红头潜鸭(Aythya ferina,易危,VU). 属于江苏省重点保护鸟类的越冬水鸟36种;属于中日候鸟保护协定的越冬水鸟33种;属于中澳候鸟保护协定的越冬水鸟5种.
2.2 水鸟多样性基于近3年调查数据分析显示,石臼湖越冬水鸟数量、物种数、Shannon-Wiener多样性指数和Inverse-Simpson多样性指数均最高(表 2);金牛湖和长荡湖越冬水鸟数量最少,且物种数也最低,均为11种;Shannon-Wiener多样性指数最低的是长荡湖,Inverse-Simpson多样性指数最低的是昆承湖(表 2).
| 表 2 江苏沿江代表性湖泊湿地越冬水鸟数量、物种数和多样性指数* Tab. 2 Wintering waterbirds abundance, number of species and diversity indices in lake wetlands of Yangtze River coast in Jiangsu Province |
江苏沿江代表性湖泊湿地间越冬水鸟β多样性如表 3. 其中,石臼湖与其它湿地间的β多样性最大,均大于0.6;长荡湖水鸟群落与其它湿地间的相异性也较高,其它湿地间的β多样性相对较小.
| 表 3 江苏沿江代表性湖泊湿地间越冬水鸟β多样性指数 Tab. 3 Wintering waterbirds β diversity indices among lake wetlands of Yangtze River coast in Jiangsu Province |
基于Sørensen的越冬水鸟总体β多样性指数(βsor)为0.679,其中物种空间周转组分(βsim)为0.520,嵌套组分(βsne)为0.160,表明物种替换是该区域β多样性形成的主要原因. 石臼湖对区域β多样性的相对贡献显著高于其它湿地,其次是长荡湖,相对贡献最低的是尚湖,其它湿地对区域β多样性的相对贡献相近(图 2).

|
图 2 江苏沿江10处湖泊湿地对区域β多样性的相对贡献(**表示石臼湖对区域β多样性有显著贡献) Fig.2 Local contributions to β diversity of lake wetlands along the Yangtze River coast in Jiangsu Province |
越冬水鸟丰富度与湖泊湿地总面积呈显著正相关,与缓冲区植被和水体面积呈负相关. 人均GDP与越冬水鸟丰富度呈显著负相关;越冬水鸟物种数与湖泊湿地总面积呈正相关,与人均GDP呈负相关;湖泊湿地总面积、缓冲区植被、水体面积以及人均GDP对Shannon-Wiener多样性指数和Inverse-Simpson多样性指数的影响均不显著(表 4).
| 表 4 基于线性模型的参数估计分析不同环境和人为因素对越冬水鸟多样性和丰富度的影响 Tab. 4 Results of the parameter estimated of environmental and anthropogenic factors which affecting wintering waterbirds diversity and abundance |
长江中下游流域是全球水鸟多样性的热点区域,历来受到相关学者的广泛关注[14, 17]. 位于长江下游的江苏沿江地区经济相对发达,其湿地水鸟种类、丰富度和多样性与上游地区存在一定差异,然而目前对该区域越冬水鸟的研究还相对较少,且多集中于单个湿地[26-27]. 本研究以江苏长江沿江代表性湖泊湿地为对象,基于近3年(2018-2020年)越冬水鸟调查数据,分析该区域越冬水鸟的群落组成和多样性,结合环境和人为因素,阐明区域越冬水鸟丰富度和多样性的影响因素,有利于长江中下游流域越冬水鸟的全面保护. 同时,随着长江经济带的飞速发展,流域其它地区也或将面临相似的保护困境[28],因此,基于对较发达的江苏沿江湖泊湿地越冬水鸟多样性的研究,也可为流域其它地区的保护工作提供现实参照.
3.1 湿地越冬水鸟的种类和丰富度2018-2020年间共记录越冬水鸟42种,包括多种国家级重点保护物种,如东方白鹳、小天鹅、白琵鹭等. 区域越冬水鸟物种数与安徽升金湖国家级自然保护区2015年记录的44种相当[29],虽与鄱阳湖区2016年记录到的68种存在一定的差距[30],但多于2019年西洞庭湖的33种[31],显示了该区域较高的物种多样性.
2019和2020年的调查中,10处代表性湖泊湿地越冬水鸟总数均达到4.5万只左右,以鸭科鸟类为主. 石臼湖2020年越冬水鸟25种,数量达2万只. 1980s石臼湖共记录到越冬水鸟33种,包括大鸨、丹顶鹤、黄嘴白鹭等国家级保护物种[32],然而现在已难觅其踪迹. 调查记录到小天鹅最大数量约8200只,远超国际重要湿地标准9,即1 % 标准. 2005年长江中下游越冬水鸟同步调查中,江苏全省未见小天鹅分布[33],2011和2015年石臼湖小天鹅的数量为3057只[34-35],可见其数量有大幅增长;太湖越冬水鸟的数量也相对较多,主要类群为鸭科鸟类和鸥科鸟类,但由于缺乏历史数据,其鸟类群落结构和数量变化尚无法评估;赤山湖、尚湖等也有较多的鸭科鸟类分布. 其中,雍凡等[26]于2013-2018年对赤山湖越冬鸟类种群动态开展了研究,发现冬季最高越冬水鸟种类为18种,且鸭科鸟类有所增加,鸻鹬类有减少的趋势,这一结果也与长江中下游流域越冬水鸟群落结构类似[17].
3.2 湿地越冬水鸟的多样性指数石臼湖Shannon-Wiener多样性指数和Inverse-Simpson多样性指数均最高,长荡湖则相对较低,其它湖泊的多样性指数差异不大,这一结果可能与其环境有关,将在下文展开讨论. β多样性分析结果显示,石臼湖越冬水鸟群落与其它湿地间差异最大,长荡湖次之,这一结果与上述α多样性结果一致.
β多样性分解结果表明,物种空间周转组分(βsim)远高于嵌套组分(βsne),表明物种周转是该区域β多样性形成的主要原因,不同物种趋于分布在不同的湿地中. 环境过滤是导致物种周转的潜在机制之一[36],调查发现石臼湖越冬水鸟种类最多,其对区域β多样性的贡献也显著高于其它湿地,可能是其栖息地异质性较高,有利于不同物种的生存,因而也具有较高的保护价值[37]. 长荡湖对区域β多样性的贡献也相对较高,可能由于其物种数较少,物种的消失降低了与其它湿地的群落相似性,进而增加了其对β多样性的贡献[38].
3.3 环境和人为因素对越冬水鸟多样性和丰富度的影响湖泊湿地面积与越冬水鸟物种数和丰富度呈显著正相关,这一结果与岛屿生物地理学等理论相符合[27, 39]. 较大的湿地可能提供更多样化的觅食场所和食物来源[40-41],同时可以降低种间竞争的压力[42]和捕食风险[43],因而可以吸引更多种类的水鸟来此越冬. 石臼湖越冬水鸟多样性和丰富度均最高,可能与其湖泊特征有关,湿地的平均坡度可能是影响鸟类丰富度的重要因素[44]. 石臼湖是研究区唯一的“蝶形”湖泊,水文环境的变化形成了大量草洲和泥滩,这种多样化的栖息地类型对其水鸟多样性和丰富度的维持具有重要意义.
江苏是长江中下游地区经济最发达的区域,有关其沿江湖泊湿地越冬水鸟多样性及其影响因素的研究仍然相对缺乏[45]. 与上游湿地不同,江苏沿江湖泊湿地除石臼湖外,其它湿地水文环境的变化对湿地景观的影响较小,很少形成草洲、泥滩等栖息地类型,人为干扰可能是影响其鸟类多样性的主要因素. 本研究结果显示,湿地所在地区GDP与水鸟丰富度和物种数均显著负相关,表明人类活动对其影响显著,这也与前人相关研究结果相符[46-47]. 研究区所在的江苏苏南地区是全国经济最发达、人口最稠密的区域之一,人类活动如工农业生产、城市化等过程可能对越冬水鸟物种数和丰富度产生直接或间接的负面影响[19, 48],例如由人类活动产生的“恐惧景观”间接降低候鸟对湿地的利用率. 湖泊湿地面积与人均GDP对越冬水鸟Shannon-Wiener和Inverse-Simpson多样性指数的影响均不显著,可能是由于其不仅取决于物种数,也与物种的个体数目有关,部分鸟类如东方白鹳、白额雁等仅分布在少数湖泊湿地且数量较少,影响了分析结果.
有研究表明,保护投入与生物多样性保护效率呈正相关[49]. 环境库兹涅茨曲线(Environmental Kuznets Curve)亦预测,当经济发展达到一个临界点时,经济发展将有助于环境的改善,进而可能吸引更多的鸟类[50]. 近年来,江苏沿江各市加大了对生物多样性保护的投入,积极开展野生动物保护和湿地修复工作,取得了一定的成效,如调查中发现东太湖近两年已经吸引了一定数量的小天鹅越冬,这也可能与苏州市近年来较高的保护投入有关.
4 保护建议人类和水鸟都依赖湿地生存[51],实现经济发展与环境保护的良性互动,是我们当前面临的重要课题. 江苏苏南地区经济发达,人类活动强度高,但也支持了较多的水鸟在此越冬,因此需采取措施以更好地维护水鸟的越冬安全. 基于本研究结果,对区域越冬水鸟保护提出如下建议:
1) 加强该区域越冬水鸟监测工作,及时发现保护空缺,对重点湖泊湿地采取针对性的保护措施. 例如在石臼湖,目前江苏境内仍没有建立正式的保护机构,安徽境内也仅为省级保护区,当前的保护效率相对较低,人为干扰强度较高,不利于越冬水鸟的保护. 因此建议将石臼湖作为苏南地区候鸟保护优先区域,设置高等级的保护区,对越冬水鸟开展重点保护.
2) 用更严格的措施保护湿地资源,减少人类活动对湿地的侵占和干扰. 2016年,江苏省出台了《江苏省湿地保护条例》,为全省湿地资源保护提供了依据,然而,苏南地区人类活动强度高,人口密度大,城市化、工业化的发展仍不时侵占自然湿地,导致湿地面积萎缩和破碎化,直接或间接影响越冬水鸟栖息地数量和质量,导致湿地生命力降低,生态功能弱化. 《中华人民共和国湿地保护法》已经实施,在将来开展土地利用规划时,需严格遵守各项相关法律、法规的规定,坚决打击和处理侵占和破坏湿地的行为;同时,对于鸟类资源较为丰富的湿地,需划定一定范围的缓冲区并加以保护,为水鸟提供较为稳定的越冬环境.
3) 安排充足的财政经费,开展有效的湿地保护和修复工程,吸引更多的水鸟来此越冬. 充足且稳定的经费投入是开展有效保护的保障,多年来,区域各级政府已经投入了大量的人力、物力和财力开展湿地保护和修复工作,取得了一定的成效. 在后续工作中,尤其需保持稳定、持续的经费投入,积极营造越冬水鸟适宜栖息地,维持并提升保护成效.
5 附录附图Ⅰ和附表Ⅰ~Ⅱ见电子版(DOI:10.18307/2022.0616).

|
附图Ⅰ 江苏沿江10处代表性湿地越冬水鸟调查位点分布 AttachedFig1 The survey points design of the 10 typical wetlands of the Yangtze coast for wintering birds in Jiangsu Province |
| 附表Ⅰ 江苏沿江10处代表性湿地越冬水鸟调查位点分布 Appendix Tab.Ⅰ A sample of the record sheet for wintering waterbirds survey |
| 附表Ⅱ 2018-2020年江苏沿江10处典型湿地越冬水鸟物种名录 Appendix Ⅱ List of the wintering waterbirds recorded in 10 typical wetlands along the Yangtze River coast in Jiangsu Province during 2018-2020 |
| [1] |
Xu WH, Fan XY, Ma JG et al. Hidden loss of wetlands in China. Current Biology, 2019, 29(18): 3065-3071. DOI:10.1016/j.cub.2019.07.053 |
| [2] |
Zedler JB, Kercher S. Wetland resources: Status, trends, ecosystem services, and restorability. Annual Review of Environment and Resources, 2005, 30: 39-74. DOI:10.1146/annurev.energy.30.050504.144248 |
| [3] |
Xu XB, Jiang B, Tan Y et al. Lake-wetland ecosystem services modeling and valuation: Progress, gaps and future directions. Ecosystem Services, 2018, 33: 19-28. DOI:10.1016/j.ecoser.2018.08.001 |
| [4] |
Song W, Deng XZ. Land-use/land-cover change and ecosystem service provision in China. Science of the Total Environment, 2017, 576: 705-719. DOI:10.1016/j.scitotenv.2016.07.078 |
| [5] |
Dong YF, Zheng WX, Zhang CX et al. Temporal and spatial differences of lake ecosystem regime shift in China. J Lake Sci, 2021, 33(4): 992-1003. [董一凡, 郑文秀, 张晨雪等. 中国湖泊生态系统突变时空差异. 湖泊科学, 2021, 33(4): 992-1003. DOI:10.18307/2021.0403] |
| [6] |
Brandis KJ, Bino G, Spencer JA et al. Decline in colonial waterbird breeding highlights loss of Ramsar wetland function. Biological Conservation, 2018, 225: 22-30. DOI:10.1016/j.biocon.2018.06.022 |
| [7] |
MacArthur RH, Wilson EO. The theory of island biogeography. Princeton: Princeton University Press, 2001: 1-7.
|
| [8] |
Gao BY, Gong P, Zhang WY et al. Multiscale effects of habitat and surrounding matrices on waterbird diversity in the Yangtze River Floodplain. Landscape Ecology, 2021, 36(1): 179-190. DOI:10.1007/s10980-020-01131-4 |
| [9] |
Zhang F, Wang M, Zhong ZF et al. The bird community in different crops periods of typical farmland and its utilization of habitats in Yancheng Nature Reserve. Ecological Science, 2019, 38(2): 36-44. [张芳, 王淼, 钟稚昉等. 盐城自然保护区典型农田不同作物期的鸟类群落及其对生境的利用. 生态科学, 2019, 38(2): 36-44.] |
| [10] |
Zhang Y, Fox AD, Cao L et al. Effects of ecological and anthropogenic factors on waterbird abundance at a Ramsar Site in the Yangtze River Floodplain. Ambio, 2019, 48(3): 293-303. DOI:10.1007/s13280-018-1076-1 |
| [11] |
Zhu XF, Lu YT, Wu PH et al. Spatial-temporal analysis of landscape ecological risk in different seasons during the past 30 years in Lake Shengjin wetland, lower reaches of the Yangtze River. J Lake Sci, 2020, 32(3): 813-825. [朱娴飞, 陆雨婷, 吴鹏海等. 近30年长江下游升金湖湿地不同季节景观生态风险时空分析. 湖泊科学, 2020, 32(3): 813-825. DOI:10.18307/2020.0321] |
| [12] |
Rösner S, Mussard-Forster E, Lorenc T et al. Recreation shapes a "landscape of fear" for a threatened forest bird species in Central Europe. Landscape Ecology, 2014, 29(1): 55-66. DOI:10.1007/s10980-013-9964-z |
| [13] |
Wu CS, Murray AT. Estimating impervious surface distribution by spectral mixture analysis. Remote Sensing of Environment, 2003, 84(4): 493-505. DOI:10.1016/S0034-4257(02)00136-0 |
| [14] |
Cao L, Meng FJ, Zhao QS. Understanding effects of large-scale development on bird migration and habitats through cutting edge avian monitoring techniques. Bulletin of Chinese Academy of Sciences, 2021, 36(4): 436-447. [曹垒, 孟凡娟, 赵青山. 基于前沿监测技术探讨"大开发"对鸟类迁徙及其栖息地的影响. 中国科学院院刊, 2021, 36(4): 436-447.] |
| [15] |
Johnson CN, Balmford A, Brook BW et al. Biodiversity losses and conservation responses in the Anthropocene. Science, 2017, 356(6335): 270-275. DOI:10.1126/science.aam9317 |
| [16] |
Leprieur F, Beauchard O, Blanchet S et al. Fish invasions in the world's river systems: When natural processes are blurred by human activities. PLoS Biology, 2008, 6(2): e28. DOI:10.1371/journal.pbio.0060028 |
| [17] |
Cao L, Zhang Y, Barter M et al. Anatidae in Eastern China during the non-breeding season: Geographical distributions and protection status. Biological Conservation, 2010, 143(3): 650-659. DOI:10.1016/j.biocon.2009.12.001 |
| [18] |
Fang JY, Wang ZH, Zhao SQ et al. Biodiversity changes in the lakes of the Central Yangtze. Frontiers in Ecology and the Environment, 2006, 4(7): 369-377. DOI:10.1890/1540-9295(2006)004[0369:BCITLO]2.0.CO;2 |
| [19] |
Xu YJ, Si YL, Yin SL et al. Species-dependent effects of habitat degradation in relation to seasonal distribution of migratory waterfowl in the East Asian-Australasian Flyway. Landscape Ecology, 2019, 34(2): 243-257. DOI:10.1007/s10980-018-00767-7 |
| [20] |
Zhou Y, Jing L, Jia YF et al. Population dynamics of four endangered cranes and landscape patterns of habitats in the East Dongting Lake during recent 30 years. J Lake Sci, 2019, 31(5): 1415-1423. [周延, 靖磊, 贾亦飞等. 东洞庭湖4种濒危鹤类近30年种群动态及栖息地景观格局变化. 湖泊科学, 2019, 31(5): 1415-1423. DOI:10.18307/2019.0515] |
| [21] |
Zheng GM. A checklist on the classification and distribution of the birds of China (Third Edition). Beijing: Science Press, 2017. [郑光美. 中国鸟类分类与分布名录. 北京: 科学出版社, 2017.]
|
| [22] |
Ma KP, Liu YM. Measurement methods of community diversityⅠ α diversity measurement methods (Ⅱ). Biodiversity Science, 1994, 2(4): 231-239. [马克平, 刘玉明. 生物群落多样性的测度方法Ⅰ α多样性的测度方法(下). 生物多样性, 1994, 2(4): 231-239.] |
| [23] |
Si XF, Zhao YH, Chen CW et al. Beta-diversity partitioning: Methods, applications and perspectives. Biodiversity Science, 2017, 25(5): 464-480. [斯幸峰, 赵郁豪, 陈传武等. Beta多样性分解: 方法、应用与展望. 生物多样性, 2017, 25(5): 464-480. DOI:10.17520/biods.2017024] |
| [24] |
Wang T, Zhou LZ. The spatial-temporal patterns of bird diversity and its determinants in the small wetlands in Hefei City. Biodiversity Science, 2022, Online. DOI: 10.17520/biods.2021445. [汪婷, 周立志. 合肥市小微湿地鸟类多样性的时空格局及其影响因素. 生物多样性, 2022, 在线发表. ]
|
| [25] |
López-Delgado EO, Winemiller KO, Villa-Navarro FA. Local environmental factors influence beta-diversity patterns of tropical fish assemblages more than spatial factors. Ecology, 2020, 101(2): e02940. DOI:10.1002/ecy.2940 |
| [26] |
Yong F, Zhang WW, Wu Y et al. Population dynamics of wintering waterbirds in Chishan Lake national wetland park in Jurong, Jiangsu Province. Wetland Science, 2019, 17(6): 651-657. [雍凡, 张文文, 吴翼等. 江苏句容赤山湖国家湿地公园越冬水鸟种群动态研究. 湿地科学, 2019, 17(6): 651-657.] |
| [27] |
Sun Y, Deng CS, Lu CH. Impact of reed cutting on bird diversity and spatial distribution in Taihu national wetland park in winter. Wetland Science, 2014, 12(6): 697-702. [孙勇, 邓昶身, 鲁长虎. 芦苇收割对太湖国家湿地公园冬季鸟类多样性和空间分布的影响. 湿地科学, 2014, 12(6): 697-702.] |
| [28] |
Sun CZ, Konig HJ, Uthes S et al. Protection effect of overwintering water bird habitat and defining the conservation priority area in Poyang Lake wetland, China. Environmental Research Letters, 2020, 15(12): 125013. DOI:10.1088/1748-9326/abc6d0 |
| [29] |
Song YW, Zhou LZ. Effects of habitat changes on spatio-temporal pattern of the wintering waterbrid community at Shengjin Lake. Journal of Anhui Agricultural University, 2019, 46(4): 610-617. [宋昀微, 周立志. 生境变化对升金湖越冬水鸟群落时空动态的影响. 安徽农业大学学报, 2019, 46(4): 610-617.] |
| [30] |
Zhang N, Li YK, Shan JH et al. Community structure, abundance and spatial distribution of water birds wintering in Poyang Lake wetland. J Lake Sci, 2019, 31(1): 183-194. [张娜, 李言阔, 单继红等. 鄱阳湖枯水期延长背景下越冬水鸟群落结构、丰富度及其空间分布格局. 湖泊科学, 2019, 31(1): 183-194. DOI:10.18307/2019.0117] |
| [31] |
Zeng YB, Peng BY, Peng PB et al. Diversity and distribution pattern of wintering waterfowl in West Dongting Lake in winter. Hunan Forestry Science & Technology, 2020, 47(6): 37-46. [曾益波, 彭波涌, 彭平波等. 西洞庭湖越冬水鸟多样性及其空间分布. 湖南林业科技, 2020, 47(6): 37-46.] |
| [32] |
Wang QS, Hu XL, Xing QR. Waterfowls of ShiJiu Lake in Anhui Province. Journal of Anhui University: Natural Sciences, 1983, 7(1): 115-124. [王岐山, 胡小龙, 邢庆仁. 安徽石臼湖的水禽. 安徽大学学报: 自然科学版, 1983, 7(1): 115-124.] |
| [33] |
Barter M, Lei G, Cao L et al. Waterbird suevry of the middle and lower Yangtze river floodplain in February. Beijing: China Forestry Publishing House, 2005: 38-44. [马克 ·巴特, 雷刚, 曹垒等. 长江中下游水鸟调查报告(2005年2月). 北京: 中国林业出版社, 2005: 38-44.]
|
| [34] |
Lei G, Lei JY, Tao XD et al. Report on the coordinated suevey for wintering waterbird of the central and lower Yangtze in 2015. Wuhan: Wuhan office, World Wildlife Fund, 2015: 5-13. [雷刚, 雷进宇, 陶旭东等. 长江中下游水鸟调查报告2011. 武汉: 世界自然基金会武汉办公室, 2011: 5-13. ]
|
| [35] |
Tao XD, Lei JY, Richard H et al. Report on waterbird survey of the middle and lower Yangtze in 2011. Beijing: China Forestry Publishing House, 2017: 5-12. [陶旭东, 雷进宇, 理查德·赫伦等. 长江中下游水鸟调查报告(2015). 北京: 中国林业出版社, 2017: 5-12. ]
|
| [36] |
Angeler DG. Revealing a conservation challenge through partitioned long-term beta diversity: Increasing turnover and decreasing nestedness of boreal lake metacommunities. Diversity and Distributions, 2013, 19(7): 772-781. DOI:10.1111/ddi.12029 |
| [37] |
Spector S. Biogeographic crossroads as priority areas for biodiversity conservation. Conservation Biology, 2002, 16(6): 1480-1487. DOI:10.1046/j.1523-1739.2002.00573.x |
| [38] |
Tatsumi S, Iritani R, Cadotte MW. Temporal changes in spatial variation: Partitioning the extinction and colonisation components of beta diversity. Ecology Letters, 2021, 24(5): 1063-1072. DOI:10.1111/ele.13720 |
| [39] |
Connor EF, Courtney AC, Yoder JM. Individuals-area relationships: The relationship between animal population density and area. Ecology, 2000, 81(3): 734-748. DOI:10.1890/0012-9658(2000)081[0734:IARTRB]2.0.CO;2 |
| [40] |
Roslin T, Várkonyi G, Koponen M et al. Species-area relationships across four trophic levels - decreasing island size truncates food chains. Ecography, 2014, 37(5): 443-453. DOI:10.1111/j.1600-0587.2013.00218.x |
| [41] |
Elliott LH, Igl LD, Johnson DH. The relative importance of wetland area versus habitat heterogeneity for promoting species richness and abundance of wetland birds in the Prairie Pothole Region, USA. The Condor, 2019, 122(1): duz060. DOI:10.1093/condor/duz060 |
| [42] |
Buchmann CM, Schurr FM, Nathan R et al. Habitat loss and fragmentation affecting mammal and bird communities—The role of interspecific competition and individual space use. Ecological Informatics, 2013, 14: 90-98. DOI:10.1016/j.ecoinf.2012.11.015 |
| [43] |
Whittingham MJ, Evans KL. The effects of habitat structure on predation risk of birds in agricultural landscapes. Ibis, 2004, 146: 210-220. DOI:10.1111/j.1474-919X.2004.00370.x |
| [44] |
Zhang Y, Jia Q, Prins HHT et al. Effect of conservation efforts and ecological variables on waterbird population sizes in wetlands of the Yangtze River. Scientific Reports, 2015, 5: 17136. DOI:10.1038/srep17136 |
| [45] |
Xia SX, Yu XB, Lei JY et al. Priority sites and conservation gaps of wintering waterbirds in the Yangtze River floodplain. Journal of Geographical Sciences, 2020, 30(10): 1617-1632. DOI:10.1007/s11442-020-1803-y |
| [46] |
Chen S, Zhang Y, Borzée A et al. Landscape attributes best explain the population trend of wintering greater white-fronted goose (Anser albifrons) in the Yangtze River floodplain. Land, 2021, 10(8): 865. DOI:10.3390/land10080865 |
| [47] |
Cristaldi MA, Giraudo AR, Arzamendia V et al. Urbanization impacts on the trophic guild composition of bird communities. Journal of Natural History, 2017, 51(39/40): 2385-2404. DOI:10.1080/00222933.2017.1371803 |
| [48] |
Cao YN, Kong LQ, Zhang LF et al. The balance between economic development and ecosystem service value in the process of land urbanization: A case study of China's land urbanization from 2000 to 2015. Land Use Policy, 2021, 108: 105536. DOI:10.1016/j.landusepol.2021.105536 |
| [49] |
Waldron A, Miller DC, Redding D et al. Reductions in global biodiversity loss predicted from conservation spending. Nature, 2017, 551(7680): 364-367. DOI:10.1038/nature24295 |
| [50] |
Lantz V, Martínez-Espiñeira R. Testing the environmental Kuznets curve hypothesis with bird populations as habitat-specific environmental indicators: Evidence from Canada. Conservation Biology: the Journal of the Society for Conservation Biology, 2008, 22(2): 428-438. DOI:10.1111/j.1523-1739.2008.00885.x |
| [51] |
Cao L, Fox AD. Birds and people both depend on China's wetlands. Nature, 2009, 460(7252): 173. DOI:10.1038/460173b |