 2023, Vol. 35
2023, Vol. 35 


(2: 中国科学院大学, 北京 100049)
(3: 安徽师范大学, 皖江流域退化生态系统的恢复与重建省部共建协同创新中心, 芜湖 241002)
(2: University of Chinese Academy of Sciences, Beijing 100049, P. R. China)
(3: Collaborative Innovation Center of Recovery and Reconstruction of Degraded Ecosystem in Wanjiang Basin Co-founded by Anhui Province and Ministry of Education, Anhui Normal University, Wuhu 241002, P. R. China)
大型底栖无脊椎动物(以下简称底栖动物)生活在水与底质的交界面,是水生态系统的重要组成部分,其生物扰动和生理活动影响着有机质的分解,具有调节泥-水界面物质交换和促进水体自净等生态功能,同时底栖动物提供了底层生产力,直接参与生态系统的能量流动与物质循环[1-3],对维持水生态系统完整性有重要作用[4]。此外,底栖动物分布广泛但移动能力较弱,生活场所固定且生命周期较长,不同类群耐受和敏感程度差异大,其变化能反映出环境胁迫的长期变化和累积效应,是理想的指示生物[5-7]。目前,底栖动物已成为水生态系统健康评价和管理的重要指标,欧盟水框架指令及美国环境保护署发布的有关生态评估指导文件中均采用了底栖动物相关指标[7-8]。我国底栖动物生态学研究起步相对较晚,始于1960s,并随着1970s环境污染调查而逐步发展[9],“十一五”以来,在国家水体污染控制与治理科技重大专项等科技项目的支持下,大量水生生物监测与评价工作陆续在松花江、辽河、海河、淮河、长江、太湖、珠江等重点流域开展,为支撑水环境管理方式的转变奠定了基础[10-11]。然而,由于缺乏历史资料,关于底栖动物群落结构长期演变的研究较少,制约了水生态评价和管理中参照状态的确定[12-13]。
太湖是我国第三大淡水湖,地处长三角经济发达区域,是无锡、苏州、上海等城市的重要饮用水水源地,并承担着防洪、生态维护等多种生态功能[14]。1980s以来太湖的富营养化和蓝藻水华问题逐渐凸显,1990s始其治理得到重视,是国家重点治理的“三湖”之一[15]。梅梁湾位于太湖北部,紧邻无锡市,由于工农业污水不断排入湾内, 1970年后西北沿岸既见条带状分布水华[16];1990s水体富营养化加重,夏秋季大量蓝藻严重影响周边自来水厂正常制水[17];1998年全湖实施“零点行动”,但收效甚微,梅梁湾各项营养盐指标在1999年的监测中不减反增[18];2007年太湖北部蓝藻水华积聚造成饮用水危机事件[19],此后国家和地方持续提高湖泊富营养化和蓝藻水华治理力度,实施了一系列环境治理和生态修复工程。随着区域内社会经济发展影响和治理工作的不断深入,目前已有许多研究表明,太湖水环境发生了较大变化。近30年来,水体中总氮浓度呈现先上升后下降的波动变化趋势[20];2003年以来,总磷平均浓度及波动性相较2000年以前均明显下降,但在2017年后有反弹趋势[19, 21];2007年以来,与蓝藻水华紧密相关的叶绿素呈波动上升趋势,目前水平已远高于2000年以前[22]。水质变化直接影响到水生生物群落的演变,然而,当前关于太湖水生生物演变的研究较为匮乏,已有研究主要关注藻类水华、浮游植物、水生植物[15, 23-24],缺少对底栖动物演变的系统分析。
本文以太湖梅梁湾为研究对象,基于2007—2019年长期定点监测数据,结合历史文献资料,分析近40年来底栖群落结构及演变特征,并结合水体和底质理化数据,应用非度量多维尺度分析(non-metric multi dimensional scaling,NMDS)和随机森林(random forest,RF)等方法探究底栖群落演变关键驱动因子。本文旨在加深人为干扰下富营养湖泊生态环境演变规律认识,以期为湖泊水生态管理提供科学参考。
1 材料与方法 1.1 研究区域梅梁湾位于太湖北部,面积约132 km2,平均水深约2 m,西部有两条入湖河流-武进港和直湖港,污染类型主要为工农业污染,北部有一条入湖河流——梁溪河,主要为城镇生活污染[25]。中国科学院太湖湖泊生态系统研究站在梅梁湾共布设7个长期监测点位,其中TH0、TH6位于河口,其他点位位于敞水区。
1.2 常规监测底栖动物常规监测时段为2007—2019年,每季度一次,分别为每年的2、5、8、11月。样品定量采集用1/40 m2改良彼得森采泥器,每个点采集2斗混合为1个样品。泥样现场经250 μm尼龙筛洗净后,剩余物带回实验室分拣,在白瓷盘中逐一将底栖动物拣出,用7% 福尔马林溶液保存,利用解剖镜和显微镜将标本鉴定至尽可能低的分类单元[26-28]。

|
图 1 太湖梅梁湾底栖动物常规监测点 Fig.1 Routine monitoring site of macrozoobenthos in Meiliang Bay of Lake Taihu |
在采集底栖动物样品时,同时现场测定环境指标,并采集水样分析化学参数。现场用多参数水质监测仪测定pH、电导率,用塞氏盘测定透明度(SD),并采集表层和底层混合水样冷藏带回太湖湖泊生态系统研究站,分析水体高锰酸盐指数(CODMn)、总氮(TN)、硝态氮(NO3--N)、氨氮(NH3-N)、总磷(TP)、磷酸根(PO43--P)及叶绿素a(Chl.a),并计算湖泊综合营养状态指数(TLI(∑));在每年的5月份采样同时用改良彼得森采泥器采取表层沉积物样品并带回实验室测定底质总氮(TNs)含量、总磷(TPs)含量和烧失量(LOI),理化指标分析、计算方法参考《湖泊富营养化调查规范》[29]、《水和废水监测分析方法》[30]。
1.3 历史数据收集在常规监测数据基础上,为分析梅梁湾底栖动物演变特征,通过查阅期刊论文、专著,整理1980s以来底栖动物、水质、底质理化数据。其中,1980s底栖动物数据来源于《太湖环境质量调查研究》[31]及《太湖水环境及其污染控制》[32],1990s来源于《太湖环境生态研究》[33],形成历史数据集,主要包括种类组成和丰度,1960s仅有物种名录数据,在定量分析中未作考虑。在整理数据时,对物种名进行核实,合并同种异名。1980s、1990s水质和底质理化因子数据来源与同年代底栖动物相同。使用数据时,1980s、1990s数据整理后均使用该指标年均值。
1.4 数据分析 1.4.1 物种优势度使用优势度来表征不同物种在群落中的相对重要性,优势度计算方式如下:
| $ Y=\frac{n_i}{N} \cdot f_i $ | (1) |
式中, ni为站位中i种的个数;N为站位中底栖动物总个数;fi为i种在各站位中出现的频率,物种优势度Y>0.02的物种为优势种。
1.4.2 非度量多维尺度分析应用NMDS分析探究底栖动物群落演变特征,同时使用环境因子进行耦合,确定与群落演变显著相关的因子。NMDS分析基于Bray-Curtis相似性矩阵,选取两轴或者三轴使得NMDS分析应力值低于0.2,分析软件使用R语言Vegan包[34]。
1.4.3 随机森林随机森林模型(RF)是一种通过聚集大量决策树来提高预测准确性的组合模型,可用于处理非线性关系、分类、回归、高阶相关、评估变量的重要性等[35]。已有研究表明其探究生物与环境因子间关系的方面拥有优异的效果[36-37]。
本文用RF对梅梁湾底栖群落数据和环境数据的耦合,对环境因子重要性排序,得出各底栖类群的主要影响因子。RF分析通过Python语言Sklearn库实现,采用bootstrap作为组合方法以避免过拟合及噪声干扰,用GridSearchCV工具进行模型优化[38],最终得到最合适的子树数目与最佳深度,本研究中最合适子树数目结果为1000,最佳深度为10。
2 结果与分析 2.1 环境因子变化1980s以来梅梁湾的多项水体及底质理化因子变化明显(附图Ⅰ)。水体透明度年际间波动较大,介于0.20~0.78 m,均值为0.39 m;多年CODMn介于3.13~10.36 mg/L之间,均值为5.69 mg/L,2007年前后直湖港河口监测点位(TH6)CODMn异常升高,随后下降明显,反映入湖水质的改善。水体中TN、NO3--N、NH3-N浓度均在2007年前后达到峰值,并远高于1980s、1990s水平,2007年后出现大幅下降。其中,TN浓度最高值出现在2007年5月,达7.92 mg/L,时值太湖水危机事件,蓝藻大面积暴发,水质状况差,随后TN浓度逐渐下降,趋于平稳,近5年来均值为2.58 mg/L。水体中TP以2007和2010年为拐点,1980s-2007年上升明显,2007年前后到达高值(0.24 mg/L),2007—2010年略有下降,随后出现反弹,并在2017、2019年达到高值。Chl.a浓度总体呈波动上升趋势,并分别在2008、2012、2017年出现高值,2017年后波动剧烈,2017—2019年均浓度为58 μg/L。结合各项因子计算得到梅梁湾TLI指数变化,1980s湖区整体处于贫营养状态,1990s以后TLI指数均保持在较高水平,介于51.6~75.9,大多数时间处于轻度、中度富营养状态。
底质理化性质中,TNs含量在1980s、1990s总体稳定,低于1000 mg/kg,2007年(1110.71 mg/kg)至2013年呈上升趋势,随后逐渐下降,但仍高于20世纪。TPs含量在1980s、1990s较为平稳,2007年后波呈波动下降趋势。底质LOI自2007年以后明显高于1980s、1990s水平,波动较大,2007—2019年均值为4.62%。
2.2 底栖动物群落结构变化 2.2.1 物种数与丰度1980—2019年梅梁湾共记录底栖动物3门7纲43属(种),其中环节动物门11种,节肢动物门20种,软体动物门12种,各年份物种数变化范围在10~39之间,多年平均物种数24种。
底栖动物总丰度变化明显(图 2),在1980—1990s,呈缓慢上升趋势,1980年为63.1 ind./m2,至1994年达1163.1 ind./m2,优势类群为双壳纲和甲壳纲;2007年达到峰值,湖区均值丰度达7142.8 ind./m2,以耐污类群寡毛纲占绝对优势(87.2%)。2007—2017年,丰度呈先降低后上升趋势,在2012—2014年间出现低值,3年均值仅为952.9 ind./m2,主要是寡毛纲丰度下降导致,随后出现回升,2017—2019年波动剧烈,2018年均值仅为884.9 ind./m2,2019年则达3499.9 ind./m2。

|
图 2 1980—2019年梅梁湾底栖动物总丰度变化 Fig.2 Changes in total macrozoobenthos abundance in Meiliang Bay from 1980 to 2019 |
底栖动物主要类群变化明显(图 3),寡毛纲丰度在2007年出现峰值6347.1 ind./m2,2007—2015年波动下降,2015年后年际变化剧烈,整体呈上升趋势。2007年之后,总丰度变化与寡毛纲丰度变化趋势相似,主导了总丰度的变化。多毛纲在1980、1990s丰度较低,2007—2011年逐步上升,达到294.3 ind./m2,之后逐步下降。昆虫纲丰度在1980、1990s呈现缓慢上升的趋势,2007年以来波动幅度大,年均丰度介于59.3~1066.4 ind./m2,高值出现在2017年2月,达3008.6 ind./m2。腹足纲丰度在1980s、1990s呈上升趋势,2007年后总体波动下降。甲壳纲丰度在1980s、1990s水平较低,2007年后总体丰度较高,变化幅度较大,丰度介于5.71~1200 ind./m2。双壳纲丰度在20世纪变化波动大,丰度水平高(均值235.5 ind./m2),现阶段丰度处于较低水平,均值为18.9 ind./m2。

|
图 3 1980—2019年梅梁湾底栖动物各纲丰度变化 Fig.3 Changes in abundance for each taxonomic class in Meiliang Bay from 1980 to 2019 |
1980年以来,梅梁湾底栖动物群落组成发生了显著变化(图 4),2007年以来以寡毛纲占优,占比呈现先降后升的趋势,在2007和2019年占比高于90%。腹足纲与双壳纲在1980s-1990s均为优势类群,2007年后腹足纲占比极少,均未超过3%,双壳纲介于1% ~20% 之间,甲壳纲在2008—2011、2013—2014年两个阶段为优势类群,昆虫纲在全时段均有出现,2012—2018年占比较高,介于19% ~42% 之间。

|
图 4 1980—2019年梅梁湾底栖动物群落相对丰度变化 Fig.4 Changes in macrozoobenthic community relative abundance in Meiliang Bay from 1980 to 2019 |
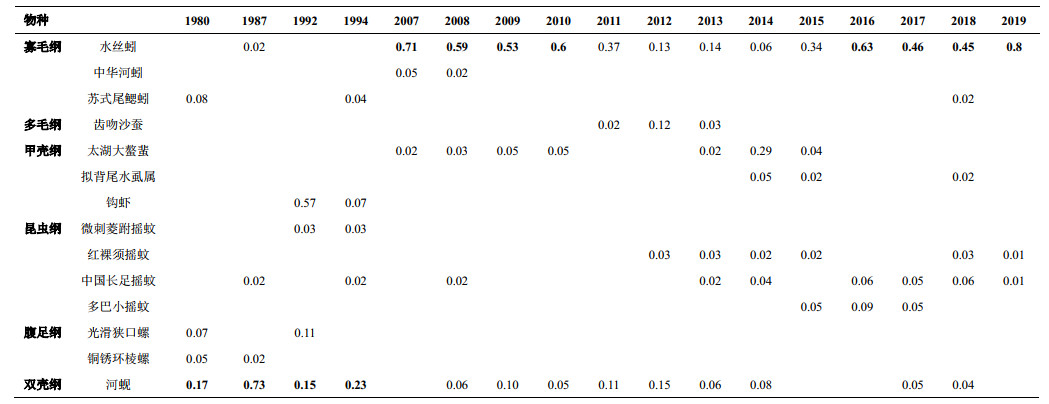
优势种分析结果显示,近40年来梅梁湾底栖动物优势种发生明显变化,1980s-1990s以较清洁的河蚬优势度最高,其次还有光滑狭口螺(Stenothyra glabra)、钩虾(Gammarus sp.)和摇蚊幼虫等作为优势种。2007—2019年几乎都以耐污的水丝蚓(Limnodrilus spp.)优势度最高,其优势度有一个先降后升的趋势,次优势种多为摇蚊幼虫(Chironomidae)、河蚬、太湖大螯蜚(Grandidierella taihuensis)等。
2.2.3 底栖动物空间格局演变从不同年份来看,底栖动物空间格局亦在发生变化(图 5)。1980、1990s敞水区表现为双壳纲、腹足纲、甲壳纲共同占优,而耐污类群已在直湖港、梁溪河河口占据较高比例,表明已受到入湖河流污染,占比呈现向大太湖降低的趋势。2007年各样点则均呈现寡毛纲绝对占优的特征,2012年双壳纲、多毛纲占比上升,2015年后又转变为寡毛纲、昆虫纲占主导,但绝对丰度明显低于2007年。

|
图 5 1980—2019年梅梁湾底栖动物空间格局变化 Fig.5 Change in spatial patterns of macrozoobenthos in Meiliang Bay from 1980 to 2019 |
NMDS分析结果表明,40年来底栖群落发生了较大变化,优势种的变动驱动了群落结构的演变,呈现出阶段性变化(图 6)。水体TN、TP、NH3-N、CODMn、Chl.a浓度及底质TPs含量与水丝蚓和摇蚊幼虫等指示种呈明显的正相关关系,而与湖球蚬(Sphaerium lacustre)、太湖大螯蜚、齿吻沙蚕(Nephtys sp.)等多毛类、甲壳类的种类显著负相关,水体透明度与河蚬、光滑狭口螺呈正相关。

|
图 6 梅梁湾底栖动物演变与环境因子的非度量多维尺度分析 Fig.6 Nonmetric multidimensional scale analysis of macrozoobenthic succession and environmental factors in Meiliang Bay |
NMDS结果亦进一步印证了底栖动物群落结构的演变,1980s-1990s,以河蚬、光滑狭口螺及铜锈环棱螺(Bellamya aeruginosa)等软体动物为代表。21世纪以来,富营养化严重,在2007—2012年,以寡毛类的水丝蚓以及摇蚊幼虫占主导,2013—2016年前后,多毛纲的齿吻沙蚕和日本沙蚕(Nereis japonica)、双壳纲的湖球蚬、太湖大鳌蜚丰度增加,2016—2019年则与2007—2012年相似。
进一步应用随机森林分析各类群演变与环境因子的关系(图 7),结果显示对寡毛纲的变化具有主要贡献的因子有LOI、TNs及PO43--P等,对昆虫纲(主要是摇蚊幼虫)的变化具有主要贡献的因子则有TNs、TPs及pH等,说明底质的理化性质主导了底栖优势耐污类群的变化。总体上看,对各类群重要性排序前6位的环境因子中,以底质TNs、TPs、LOI以及水体Chl.a、NH3-N、PO43--P为主,均在排序前6位中出现了4次及以上。

|
图 7 环境因子对梅梁湾底栖动物各纲演变的重要性排序 Fig.7 Importance of environmental factors to the succession of each taxonomic class in Meiliang Bay |
本文结合了历史资料分析底栖动物演变,2007年以前与2007—2019年的采样工具及方式基本一致,均采用彼得森采泥器采集2次,采样面积水平相似(0.05~0.125 m2)。此外,为减少鉴定误差影响,本文将各阶段鉴定结果统一至同一分类水平,以提高结果可比性。通过对梅梁湾底栖动物种类组成、优势种、空间格局等方面的定量分析,结果显示底栖动物群落发生显著变化。结合1960s的定性资料及本文结果,可将1960s以来梅梁湾底栖动物群落演变过程总体划分为4个阶段两种模式:第1阶段为1960s-1980s,此阶段梅梁湾底栖动物详细记录数据较少,但据《太湖综合调查初步报告》[39]显示,1960s全太湖共发现40种底栖动物,物种多样性丰富,以河蚬等大型软体动物为优势种,环节动物和水生昆虫次之,水体清澈,有一定的沉水植物,相应地,水体氮磷浓度除河口一带稍高,其余水域都处于较低水平,如硝态氮浓度仅为0.05~0.09 mg/L,直至1980s底栖群落仍以河蚬、环棱螺等较清洁类群占优,这一时期人类活动对水生生境干扰较弱;第2阶段为1990s-2007年,1990s开始太湖流域乡镇企业的发展致使大量污水排入太湖,水质条件恶化,至21世纪初水质类别较20世纪已普遍降低了1~2类[40],底栖生境遭到严重破坏,直湖港河口呈现明显的受污染群落特征,底栖动物群落快速退化,以耐污种占优,物种多样性减少,大型软体动物基本消失,耐污种水丝蚓丰度急剧上升,蓝藻水华暴发严重,鱼类出现小型化特征。第3阶段为2008—2016年,2007年水危机发生后,国家和地方进一步深化太湖治理工作,实施了流域污染减排、调水引流、蓝藻打捞、生态清淤、生态修复等系列工程[41],这也使得太湖水文、水质、底质及生物群落发生变化,水体总氮指标在这一时期也显著下降[20],水质呈好转趋势,梅梁湾水质、底质条件得以改善,此阶段双壳纲、甲壳纲生物略有回升,可能与富营养区域进行的生态清淤等相关措施有关,表明水生态呈恢复趋势[42],此阶段群落结构模式与第1阶段部分相似,但底栖群落仍以耐污种占优,物种多样性不高,水环境质量差于1960s-1980s。第4阶段为2017—2019年,此阶段以水丝蚓为代表的耐污种优势度和丰度反弹,生物群落多样性减少,一方面,梅梁湾的底泥疏浚工作于2013年前后完成[43],污染湖泊的底泥疏浚一般在初期对内源负荷的控制效果较明显,根据环境的不同,效果持续时间不一,但随着时间推移,均会出现反弹现象[44],另一方面,近年来太湖的蓝藻水华亦呈现加剧趋势,2017年水华强度和水体总磷浓度双双反弹[45],富营养化程度居高不下,而其中梅梁湾湖区的情况更为严重[46],上述两方面原因可能引起近几年底栖群落结构退化。4个阶段的变化也与梅梁湾营养状态的变化相符,总体而言,梅梁湾底栖动物群落结构特征的变化,正是多年来太湖生态系统对人类活动影响下富营养化进程的表征。
3.2 群落演变关键驱动因素综合NMDS和随机森林分析结果综合来看,影响梅梁湾底栖动物群落结构变化的最主要环境因子为底质总氮、总磷、烧失量及水体营养盐等指标。底栖动物长期生活于水体底部,因此底质污染状况对底栖动物的种类组成、丰度和分布有直接影响。1990s开始,人为干扰加重,各类污染物进入梅梁湾,底质中氮磷及有机质含量发生剧烈波动,由于底栖动物主要以有机碎屑为食,因此底质中有机质对底栖动物有直接影响,进而使得水丝蚓、摇蚊随底质氮磷增加优势度上升。此外,研究表明,水体中较低的溶解氧会对软体动物产生严重危害[47],且可能会造成水体中河蚬的大量死亡[48],而在高营养水平的情况下,底层溶解氧不足,大大限制了河蚬等软体动物的生长。
水体营养盐、高锰酸盐指数及叶绿素a浓度与底栖动物丰度亦显著相关,太湖常年暴发的蓝藻水华,会引起水体不同形态的营养盐浓度及叶绿素在短期内迅速增加,同时影响湖泊的光照透明度,从而引起底栖动物的变化,各指标的增加伴随着底栖动物耐污种的增加与优势度提高。水体营养盐增加是富营养化的表现,富营养水体中通常出现高丰度的水丝蚓和摇蚊幼虫[49-50],同时,随着营养水平的不断增加,底栖动物群落也会呈现逐渐被小个体的耐污种类所主导,趋于简单化的演替趋势[51],由于梅梁湾富营养化问题严重,导致了大型底栖动物物种多样性降低,水丝蚓和摇蚊幼虫等小型耐污种成为优势种,富营养化过程亦伴随着底质中营养物质和有机质含量的增加,水体和底质营养状况共同决定了底栖动物群落结构。
此外,受湖泊富营养化的影响,水生植物的衰退甚至消亡也可能导致梅梁湾底栖群落结构的变化。1988年,包括梅梁湾在内的北部湖区各水域均发现不同面积的水生植物,至2014年,北部湖区植被几乎完全消失[52],沉水植物的茎叶增加了垂直方向上水生生物的栖息地面积,根部可以为底栖动物提供栖息地以及庇护所[53],对维持底栖动物多样性具有重要作用,近30年来梅梁湾沉水植物除有菹草外,苦草已经基本消亡[23],可能是导致1980s、1990s常见的腹足纲丰度急剧下降的一个重要原因。
4 结论1) 太湖梅梁湾1980—2019年共记录底栖动物3门7纲44属(种),其中环节动物门11种,节肢动物门20种,昆虫纲14种,软体动物门12种。
2) 1980s以来梅梁湾底栖动物群落结构发生显著变化,优势种由较敏感的软体动物转变为耐污的寡毛类、摇蚊类占优。太湖梅梁湾底栖动物呈现出显著的空间异质性,群落结构特征受入湖河流影响,总体格局为敞水区群落多样性高于河口。
3) 结合1960s资料,可将梅梁湾底栖动物群落演变分为4个阶段,第1阶段为1960s-1980s,物种丰富,清洁种占优;第2阶段为1990s-2007年,物种多样性减少,耐污种占优;第3阶段为2008—2016年,群落生态略有恢复,清洁种回升;第4阶段为2017—2019年,耐污种丰度反弹,群落结构退化。
4) 近40年来梅梁湾底栖动物群落结构的变化主要受底质和水体营养元素、蓝藻水华等因子的影响,此外水生植物的衰退也是重要影响因素。
5 附录附图Ⅰ和附表Ⅰ见电子版(DOI: 10.18307/2023.0217)。

|
附图Ⅰ 1980—2019年梅梁湾水体与底质理化因子变化 AttachedFig1 Changes in physicochemical parameters of water and sediment in Meiliang Bay from 1980 to 2019 |
| 附表Ⅰ 1980—2019年梅梁湾底栖动物优势种与优势度变化 Appendix Ⅰ Changes in dominant species and dominance indices of macrozoobenthos in Meiliang Bay from 1980 to 2019 |
| [1] |
Covich AP, Austen MC, BÄRlocher F et al. The role of biodiversity in the functioning of freshwater and marine benthic ecosystems. BioScience, 2004, 54(8): 767-775. DOI:10.1641/0006-3568(2004)054[0767:TROBIT]2.0.CO;2 |
| [2] |
Vaughn CC, Hakenkamp CC. The functional role of burrowing bivalves in freshwater ecosystems. Freshwater Biology, 2001, 46(11): 1431-1446. DOI:10.1046/j.1365-2427.2001.00771.x |
| [3] |
Atkinson CL, Capps KA, Rugenski AT et al. Consumer-driven nutrient dynamics in freshwater ecosystems: From individuals to ecosystems. Biological Reviews, 2017, 92(4): 2003-2023. DOI:10.1111/brv.12318 |
| [4] |
Vanni MJ. Nutrient cycling by animals in freshwater ecosystems. Annual Review of Ecology and Systematics, 2002, 33: 341-370. DOI:10.1146/annurev.ecolsys.33.010802.150519 |
| [5] |
Chen K, Chen QW, Yu HY et al. Methods and prospects of index of biological integrity used for China river ecological health assessment. China Environmental Science, 2018, 38(4): 1589-1600. [陈凯, 陈求稳, 于海燕等. 应用生物完整性指数评价我国河流的生态健康. 中国环境科学, 2018, 38(4): 1589-1600. DOI:10.19674/2018.0192] |
| [6] |
Poikane S, Johnson RK, Sandin L et al. Benthic macroinvertebrates in lake ecological assessment: A review of methods, intercalibration and practical recommendations. Science of the Total Environment, 2016, 543: 123-134. DOI:10.1016/j.scitotenv.2015.11.021 |
| [7] |
Birk S, Bonne W, Borja A et al. Three hundred ways to assess Europe's surface waters: An almost complete overview of biological methods to implement the Water Framework Directive. Ecological Indicators, 2012, 18: 31-41. DOI:10.1016/j.ecolind.2011.10.009 |
| [8] |
Pollard AI, Hampton SE, Leech DM. The promise and potential of continental-scale limnology using the US environmental protection agency's national lakes assessment. Limnology and Oceanography Bulletin, 2018, 27(2): 36-41. DOI:10.1002/lob.10238 |
| [9] |
Yin K, Wang YY, Xu RJ et al. Constitution and development of national river basin water environmental biomonitoring system. Environmental Monitoring in China, 2014, 30(5): 114-120. [阴琨, 王业耀, 许人骥等. 中国流域水环境生物监测体系构成和发展. 中国环境监测, 2014, 30(5): 114-120. DOI:10.19316/2014.05.025] |
| [10] |
Zhang Y, Liu L, Cai YJ et al. Benthic macroinvertebrate community structure in rivers and streams of Lake Taihu Basin and environmental determinants. China Environmental Science, 2015, 35(5): 1535-1546. [张又, 刘凌, 蔡永久等. 太湖流域河流及溪流大型底栖动物群落结构及影响因素. 中国环境科学, 2015, 35(5): 1535-1546. DOI:10.3969/2015.05.034] |
| [11] |
张远, 江源. 中国重点流域水生态系统健康评价. 北京: 科学出版社, 2019.
|
| [12] |
Morse JC, Bae YJ, Munkhjargal G et al. Freshwater biomonitoring with macroinvertebrates in east Asia. Frontiers in Ecology and the Environment, 2007, 5(1): 33-42. DOI:10.1890/1540-9295(2007)5[33:FBWMIE]2.0.CO;2 |
| [13] |
Xu YP, Wang ZJ. Developing the benchmark and reference condition for monitoring and assessment of ecological integrity of surface waters. Environmental Monitoring in China, 2018, 34(6): 1-9. [许宜平, 王子健. 水生态完整性监测评价的基准与参照状态探究. 中国环境监测, 2018, 34(6): 1-9. DOI:10.19316/2018.06.01] |
| [14] |
Gu XH, Zeng QF, Mao ZG et al. Water environment change over the period 2007-2016 and the strategy of fishery improve the water quality of Lake Taihu. J Lake Sci, 2019, 31(2): 305-318. [谷孝鸿, 曾庆飞, 毛志刚等. 太湖2007—2016十年水环境演变及"以渔改水"策略探讨. 湖泊科学, 2019, 31(2): 305-318. DOI:10.18307/2019.0201] |
| [15] |
Yang J, Zhou J, Qin BQ et al. Long-term variation characteristics of zooplankton community structure in Meiliang Bay, lake Taihu. Environmental Science, 2020, 41(3): 1246-1255. [杨佳, 周健, 秦伯强等. 太湖梅梁湾浮游动物群落结构长期变化特征(1997—2017年). 环境科学, 2020, 41(3): 1246-1255. DOI:10.13227/j.hjkx.201907054] |
| [16] |
Zhu GW, Qin BQ, Zhang YL et al. Variation and driving factors of nutrients and chlorophyll-a concentrations in northern region of Lake Taihu, China, 2005-2017. J Lake Sci, 2018, 30(2): 279-295. [朱广伟, 秦伯强, 张运林等. 2005—2017年北部太湖水体叶绿素a和营养盐变化及影响因素. 湖泊科学, 2018, 30(2): 279-295. DOI:10.18307/2018.0201] |
| [17] |
Liu YB, Gao XY. Factor cluster analysis of water quality in Meiliang Bay, Taihu Lake. J Lake Sci, 1997, 9(3): 255-260. [刘元波, 高锡芸. 太湖北部梅梁湾水域水质因子聚类. 湖泊科学, 1997, 9(3): 255-260. DOI:10.18307/1997.0310] |
| [18] |
Sun WH. Comparative analysis of water quality of Taihu lake around "zero-point action". Jiang Su Environmental Science and Technology, 2003, 16(1): 35-36. [孙卫红. "零点行动"前后太湖水质比较分析. 江苏环境科技, 2003, 16(1): 35-36. DOI:10.3969/2003.01.014] |
| [19] |
Wu HY, Jia GH, Xu B et al. Analysis of variation and driving factors of total phosphorus in Lake Taihu, 1980-2020. J Lake Sci, 2021, 33(4): 974-991. [吴浩云, 贾更华, 徐彬等. 1980年以来太湖总磷变化特征及其驱动因子分析. 湖泊科学, 2021, 33(4): 974-991. DOI:10.18307/2021.0402] |
| [20] |
Fan QH, Shen HJ, Zhang T et al. Changes of total nitrogen concentration in Taihu Lake during 1987-2016. Environmental Monitoring and Forewarning, 2017, 9(6): 8-13. [范清华, 沈红军, 张涛等. 1987—2016年太湖总氮浓度变化趋势分析. 环境监控与预警, 2017, 9(6): 8-13. DOI:10.3969/2017.06.002] |
| [21] |
Zhu GW, Qin BQ, Zhang YL et al. Fluctuation of phosphorus concentration in Lake Taihu in the past 70 years and future control strategy. J Lake Sci, 2021, 33(4): 957-973. [朱广伟, 秦伯强, 张运林等. 近70年来太湖水体磷浓度变化特征及未来控制策略. 湖泊科学, 2021, 33(4): 957-973. DOI:10.18307/2021.0401] |
| [22] |
Wu DH, Jia GH, Wu HY. Chlorophyll-a concentration variation characteristics of the algae-dominant and macrophyte-dominant areas in Lake Taihu and its driving factors, 2007-2019. J Lake Sci, 2021, 33(5): 1364-1375. [吴东浩, 贾更华, 吴浩云. 2007—2019年太湖藻型和草型湖区叶绿素a变化特征及影响因子. 湖泊科学, 2021, 33(5): 1364-1375. DOI:10.18307/2021.0506] |
| [23] |
Wang Q, Zhou XD, Luo JH et al. Remote sensing monitoring and analysis of dominant species of submerged vegetation in Taihu Lake over last 30 years. Water Resources Protection, 2016, 32(5): 123-129, 135. [王琪, 周兴东, 罗菊花等. 近30年太湖沉水植物优势种遥感监测及变化分析. 水资源保护, 2016, 32(5): 123-129, 135. DOI:10.3880/2016.05.023] |
| [24] |
Song XL, Liu ZW, Pan HK et al. Phytoplankton community structure in Meiliang Bay and Lake Wuli of Lake Taihu. J Lake Sci, 2007, 19(6): 643-651. [宋晓兰, 刘正文, 潘宏凯等. 太湖梅梁湾与五里湖浮游植物群落的比较. 湖泊科学, 2007, 19(6): 643-651. DOI:10.18307/2007.0604] |
| [25] |
Liu Q, Yuan XY, Shi ZQ et al. Study on pollution sources and degree of sediments in seven rivers flowing into Taihu Lake. Journal of Anhui Agricultural Sciences, 2012, 40(15): 8698-8701. [刘泉, 袁旭音, 时志强等. 太湖西北部入湖河道的沉积物源解析与污染程度研究. 安徽农业科学, 2012, 40(15): 8698-8701. DOI:10.13989/2012.15.041] |
| [26] |
Morse JC, Yang L, Tian L eds. Aquatic insects of China useful for monitoring water quality. Nanjing, People's Republic of China: Hohai University Press, 1994.
|
| [27] |
刘月英. 中国经济动物志: 淡水软体动物. 北京: 科学出版社, 1979.
|
| [28] |
王洪铸. 中国小蚓类研究——附中国南极长城站附近地区两新种[学位论文]. 武汉: 中国科学院水生生物研究所, 1995.
|
| [29] |
金相灿, 屠清瑛. 湖泊富营养化调查规范(第二版). 北京: 中国环境科学出版社, 1990.
|
| [30] |
国家环境保护总局《水和废水监测分析方法》编委会. 水和废水监测分析方法. 北京: 中国环境科学出版社, 1989.
|
| [31] |
太湖环境质量调查研究组. 太湖水环境质量调查研究. 1982.
|
| [32] |
黄漪平. 太湖水环境及其污染控制. 北京: 科学出版社, 2001.
|
| [33] |
蔡启铭. 太湖环境生态研究. 北京: 气象出版社, 1998.
|
| [34] |
Dixon P. VEGAN, a package of R functions for community ecology. Journal of Vegetation Science, 2003, 14(6): 927-930. DOI:10.1111/j.1654-1103.2003.tb02228.x |
| [35] |
Lam KL, Cheng WY, Su YT et al. Use of random forest analysis to quantify the importance of the structural characteristics of beta-glucans for prebiotic development. Food Hydrocolloids, 2020, 108: 106001. DOI:10.1016/j.foodhyd.2020.106001 |
| [36] |
Chen K, Yu HY, Zhang JW et al. Predictive model based multimetric index of macroinvertebrates for river health assessment. Chinese Journal of Applied Ecology, 2017, 28(6): 1993-2002. [陈凯, 于海燕, 张汲伟等. 基于底栖动物预测模型构建生物完整性指数评价河流健康. 应用生态学报, 2017, 28(6): 1993-2002. DOI:10.13287/j.1001-9332.201706.015] |
| [37] |
Ding JQ, Huang WL, Liu YC et al. Estimation of forest aboveground biomass in northwest Hunan Province based on machine learning and multi-source data. Scientia Silvae Sinicae, 2021, 57(10): 36-48. [丁家祺, 黄文丽, 刘迎春等. 基于机器学习和多源数据的湘西北森林地上生物量估测. 林业科学, 2021, 57(10): 36-48. DOI:10.11707/2021.10.04] |
| [38] |
Dong W, Huang YM, Lehane B et al. XGBoost algorithm-based prediction of concrete electrical resistivity for structural health monitoring. Automation in Construction, 2020, 114: 103155. DOI:10.1016/j.autcon.2020.103155 |
| [39] |
中国科学院南京地理研究所. 太湖综合调查初步报告. 北京: 科学出版社, 1965.
|
| [40] |
Lin ZX. Analysis of water environmental change in Taihu watershed. J Lake Sci, 2002, 14(2): 111-116. [林泽新. 太湖流域水环境变化及缘由分析. 湖泊科学, 2002, 14(2): 111-116. DOI:10.18307/2002.0203] |
| [41] |
Chen HS, Song XF, Zou GY. Comprehensive improvement of water environment and ecological restoration in Taihu Lake Basin. Advances in Science and Technology of Water Resources, 2008, 28(3): 76-79. [陈荷生, 宋祥甫, 邹国燕. 太湖流域水环境综合整治与生态修复. 水利水电科技进展, 2008, 28(3): 76-79. DOI:10.3880/2008.03.019] |
| [42] |
Wang Q, Li ZH. Study on effect of ecological dredging in Meiliang Lake in the Taihu Lake. China Harbour Engineering, 2012, 32(6): 21-23. [王琦, 李中华. 太湖梅梁湖生态疏浚工程实施效果研究. 中国港湾建设, 2012, 32(6): 21-23.] |
| [43] |
Hu XD, Zhang JH, Wu PP et al. Nitrogen and phosphorus release-based study on time-effect of eco-dredging. Water Resources and Hydropower Engineering, 2016, 47(10): 58-61, 72. [胡晓东, 张建华, 吴沛沛等. 基于氮、磷释放的太湖生态清淤时间效应研究. 水利水电技术, 2016, 47(10): 58-61, 72. DOI:10.13928/2016.10.013] |
| [44] |
Fan CX, Zhang L, Wang JJ et al. The process and mechanism of the effect of lake sediment dredging on endogenous release. Chinese Science Bulletin, 2004, 49(15): 1523-1528. [范成新, 张路, 王建军等. 湖泊底泥疏浚对内源释放影响的过程与机理. 科学通报, 2004, 49(15): 1523-1528. DOI:10.3321/2004.15.010] |
| [45] |
Qin BQ, Paerl HW, Brookes JD et al. Why Lake Taihu continues to be plagued with cyanobacterial blooms through 10 years (2007-2017) efforts. Chinese Science Bulletin, 2019, 64(6): 354-356. DOI:10.1016/j.scib.2019.02.008 |
| [46] |
Pang JL, Xu YH, He YX et al. Molecular characteristics of surface dissolved organic matter in Meiliang Bay of Lake Taihu over the algal blooming-disappearance cycle. J Lake Sci, 2020, 32(6): 1599-1609. [庞佳丽, 许燕红, 何毓新等. 太湖梅梁湾藻华暴发-消退周期表层水体溶解性有机质分子特征. 湖泊科学, 2020, 32(6): 1599-1609. DOI:10.18307/2020.0603] |
| [47] |
Saloom ME, Scot duncan R. Low dissolved oxygen levels reduce anti-predation behaviours of the freshwater clam Corbicula fluminea. Freshwater Biology, 2005, 50(7): 1233-1238. DOI:10.1111/j.1365-2427.2005.01396.x |
| [48] |
Johnson PD, McMahon RF. Effects of temperature and chronic hypoxia on survivorship of the zebra mussel (Dreissena polymorpha) and Asian clam (Corbicula fluminea). Canadian Journal of Fisheries and Aquatic Sciences, 1998, 55: 1564-1572. DOI:10.1139/F98-030 |
| [49] |
Gong ZJ, Xie P, Tang HJ et al. The influence of eutrophycation upon community structure and biodiversity of macrozoobenthos. Acta Hydrobiologica Sinica, 2001, 25(3): 210-216. [龚志军, 谢平, 唐汇涓等. 水体富营养化对大型底栖动物群落结构及多样性的影响. 水生生物学报, 2001, 25(3): 210-216. DOI:10.3321/2001.03.002] |
| [50] |
Hooper HL, Sibly RM, Hutchinson TH et al. The influence of larval density, food availability and habitat longevity on the life history and population growth rate of the midge Chironomus riparius. Oikos, 2003, 102(3): 515-524. DOI:10.1034/j.1600-0706.2003.12536.x |
| [51] |
Cai YJ, Jiang JH, Zhang L et al. Community structure and biodiversity of macrozoobenthos of typical lakes in the middle and lower reaches of the Yangtze River. J Lake Sci, 2010, 22(6): 811-819. [蔡永久, 姜加虎, 张路等. 长江中下游湖泊大型底栖动物群落结构及多样性. 湖泊科学, 2010, 22(6): 811-819. DOI:10.18307/2010.0602] |
| [52] |
Zhao K, Zhou YF, Jiang ZL et al. Changes of aquatic vegetation in Lake Taihu since 1960s. J Lake Sci, 2017, 29(2): 351-362. [赵凯, 周彦锋, 蒋兆林等. 1960年以来太湖水生植被演变. 湖泊科学, 2017, 29(2): 351-362. DOI:10.18307/2017.0211] |
| [53] |
Walker PD, Wijnhoven S, van der Velde G. Macrophyte presence and growth form influence macroinvertebrate community structure. Aquatic Botany, 2013, 104: 80-87. DOI:10.1016/j.aquabot.2012.09.003 |